当前位置:首页 > 行业资料 > 其它行业文档 > CRISPR及其在原核生物防御系统中作用的研究进展
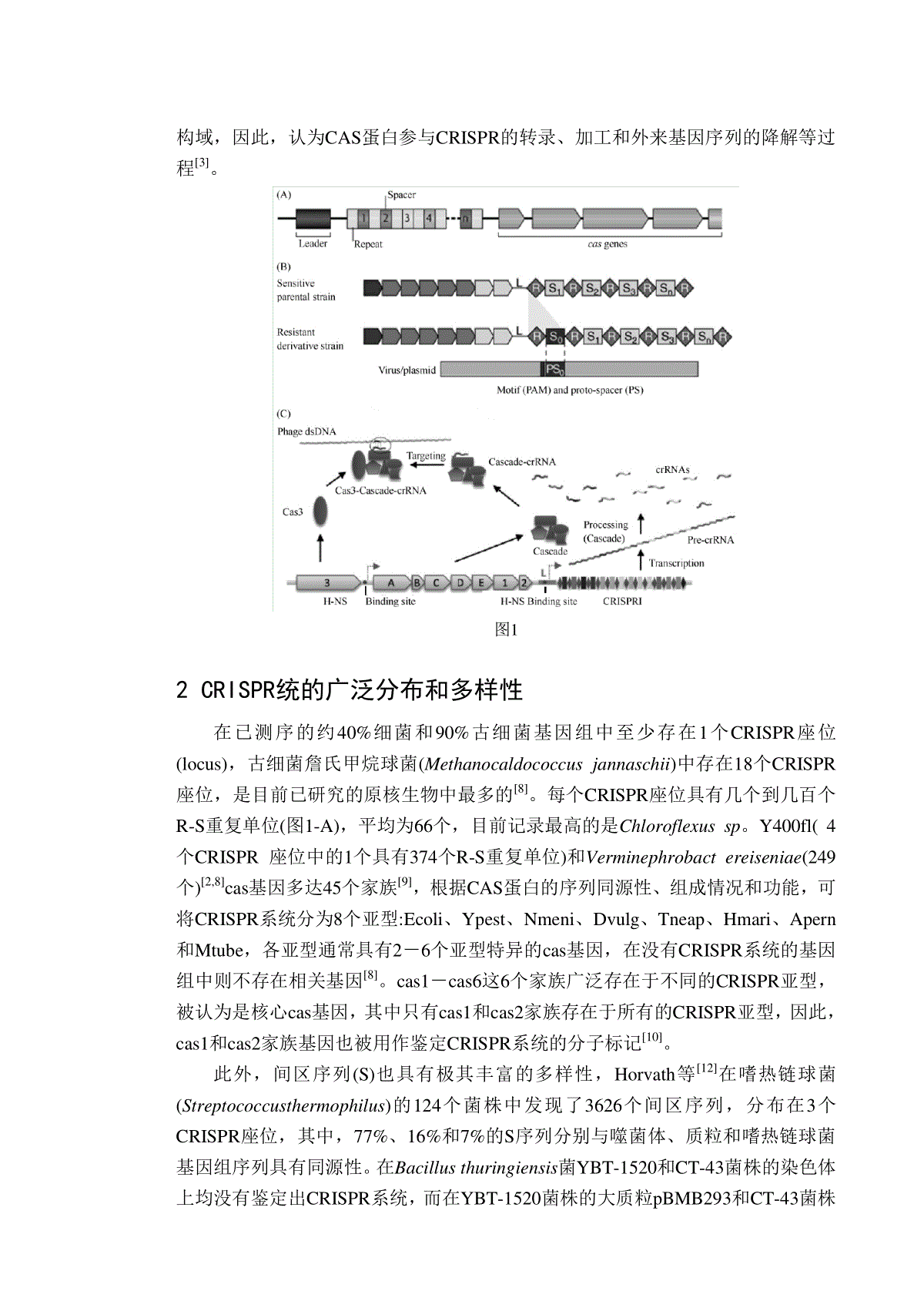
CRISPR及其在原核生物防御系统中作用的研究进展摘要近来在众多原核生物的基因组的分析中,发现一类成簇的、有规律间隔的短回文重复序列(clusteredregularlyinterspacedshortpalindromicrepeats,CRISPR)结构家族。CRISPR由一类DNA重复单元所构成,在功能上与DNA的重组、修复及原核生物的免疫防御系统关系密切。通过对于CRISPR及其相关系统(CASs)基因的比较性分析,CRISPR独特的结构和功能引起人们的广泛关注。实验表明CASs的插入序列来源于细胞内的许多染色体外遗传物质,并且以一种类似于真核生物中RNA干扰(RNAi)系统的机制,在原核生物抵抗入侵的噬菌体和质粒的防御系统中发挥着重要作用。本文综述了CRISPR系统的基本结构、多样性、作用机理及其区分自我与非我的机制,并对CRISPR研究和应用前景进行了展望。关键词:CRISPR;原核生物;免疫成簇的规律间隔的短回文重复序列(clusteredregularlyinterspacedshortpalindromicrepeats,CRISPR)是一类独特的DNA直接重复序列家族,广泛存在于众多原核生物基因(大多数的细菌和几乎所有的古细菌)中[1]。自2002年首次被人们所定义以来,CRISPR一直以其奇特的结构与特殊的功能吸引着各国科学家们的共同关注。它的结构非常稳定,长度约25~50bp的重复序列(这里我们称为repeats)被单一序列(这里我们称为spacers)间隔[2]。根据不同的研究结果,人们对CRISPR系统的生物学功能曾有过种种推测,最近人们发现Cas系统(CRISPR-associatedsequencessystem,CASs)在原核生物中表现出某种获得性免疫功能,帮助细菌抵御病毒的侵犯,并和DNA的重组与修复有关[3]。2005年,3个研究小组分别发现一些CRISPR的间区序列是来自噬菌体或质粒等染色体外的序列,据此推断CRISPR系统能使宿主获得抵抗噬菌体、质粒等外来DNA入侵的免疫能力[4-6]。2007年,Barrangou等[7]通过实验证实了CRISPR系统的这一生物学功能。1CRISPR系统的基本结构随着细菌基因组学的发展,CRISPR的基本结构已逐渐被确定,由短的高度保守的repeat与长度相似的非重复的spacers序列间隔排列[11]所组成(图1-A)。Repeats是一组高度保守的短小序列,其长度一般为25至50个碱基对。Spacers与repeats的长度相近,约为26到72个碱基对。此外一组约由4~10个保守基因组成的序列被发现于CRISPRs周围,我们称之为CASs(CRISPR-associatedsequences,CASs)[11]。在CAS蛋白中已鉴定出核酸内切酶、核酸外切酶、螺旋酶、RNA-和DNA-结合等结构域,因此,认为CAS蛋白参与CRISPR的转录、加工和外来基因序列的降解等过程[3]。图12CRISPR统的广泛分布和多样性在已测序的约40%细菌和90%古细菌基因组中至少存在1个CRISPR座位(locus),古细菌詹氏甲烷球菌(Methanocaldococcusjannaschii)中存在18个CRISPR座位,是目前已研究的原核生物中最多的[8]。每个CRISPR座位具有几个到几百个R-S重复单位(图1-A),平均为66个,目前记录最高的是Chloroflexussp。Y400fl(4个CRISPR座位中的1个具有374个R-S重复单位)和Verminephrobactereiseniae(249个)[2,8]cas基因多达45个家族[9],根据CAS蛋白的序列同源性、组成情况和功能,可将CRISPR系统分为8个亚型:Ecoli、Ypest、Nmeni、Dvulg、Tneap、Hmari、Apern和Mtube,各亚型通常具有2-6个亚型特异的cas基因,在没有CRISPR系统的基因组中则不存在相关基因[8]。cas1-cas6这6个家族广泛存在于不同的CRISPR亚型,被认为是核心cas基因,其中只有cas1和cas2家族存在于所有的CRISPR亚型,因此,cas1和cas2家族基因也被用作鉴定CRISPR系统的分子标记[10]。此外,间区序列(S)也具有极其丰富的多样性,Horvath等[12]在嗜热链球菌(Streptococcusthermophilus)的124个菌株中发现了3626个间区序列,分布在3个CRISPR座位,其中,77%、16%和7%的S序列分别与噬菌体、质粒和嗜热链球菌基因组序列具有同源性。在Bacillusthuringiensis菌YBT-1520和CT-43菌株的染色体上均没有鉴定出CRISPR系统,而在YBT-1520菌株的大质粒pBMB293和CT-43菌株的大质粒pCT283上各存在2个CRISPR座位。pBMB293的CRISPR-1座位包含10个间区序列,其中的5个分别与已知噬菌体GIL16c和Bam35c、质粒pGIL01和pAH187、HalothermothrixoreniiH168的染色体有很高的序列同源性;CRISPR-2座位包含5个间区序列,只有1个与已知的质粒序列(pAH187)具有同源性。3SpacerDNA的来源CRISPR广泛存在于原核生物(如古细菌、革兰氏阳性和革兰氏阴性细菌),并在其生理和种族发生过程中发挥着多重功能[13,14]。在来源于不同基因组的CRISPR之中,repeats所表现出的相似性是有限的,而保守基序spacers却是相同的,即使是在亲缘关系较远的古细菌和细菌中也是如此[4,15]。此前,Mojica等将这类Spacers都描述为特有的独一无二的序列,但最近的研究越来越倾向于另一种假设:CRISPR的插入序列来源于一些已存在于细胞中的序列[16]。通过检测发现,在许多株群,CRISPR中的spacers与细胞中其它遗传成分存在着高度的相似性。关于CRISPR插入片段的来源有如下几种:约65%与噬菌体和结合性质粒存在相关,其余的35%与染色体序列有关而与外缘DNA之间没有关系。之后的一系列研究使我们更清晰的了解到CRISPR的spacer元件是染色体外遗传物质入侵细菌后的残留体,他们很可能通过编码一类anti-senseRNA为细胞提供了一种对抗噬菌体感染甚至是更广泛的外源基因侵染的免疫能力[17]。4CRISPR的作用机理4.1新间区序列的获得在噬菌体等入侵宿主菌后,CRISPR系统会从外来序列中选取一段序列作为新的间区序列,加工成新的R-S序列后以非同源重组的方式增加到CRISPR簇内,从而使宿主获得抵抗相应噬菌体等再次入侵的免疫能力[8],这一现象也得到实验证实[7]。新的R-S序列几乎总插入到前导序列和原来的第一个R区之间(图1-B),因此,前导序列中可能存在相关蛋白(可能是CAS蛋白)的识别位点,在增加新R-S序列过程中起定位的功能。CRISPR系统从噬菌体等基因组中选取新的间区序列并不是随机的。事实上,多个研究小组已经在噬菌体、质粒中发现了一些称为前间区序列(protospacer)的序列,并且在其附近鉴定出一些小的前间区序列邻近基序(protospaceradjacentmotifs,PAMs)。不同CRISPR亚型识别的PAM基序具有亚型特异性[18]。目前,有哪些蛋白质参与PAM基序的识别、前间区序列的选取、间区序列长度如何确定等方面的具体机制尚不清楚。4.2CRISPR的转录与加工研究表明,CRISPR簇首先转录为长的转录体,即前crRNA,然后逐步被加工成小的crRNA。多数情况下,只有CRISPR簇的编码链被转录和加工[10]但在古细菌Sulfolobus属[19]和嗜热链球菌[7],CRISPR簇的2条链都可以被转录。在大肠杆菌K12菌株中,CRISPR簇的前crRNA与CAS蛋白CasABCDE形成的Cascade复合物结合,其中,CasE的功能是将前crRNA切割加工成crRNA。成熟的crRNA分子一般包含5'-端为8-9nt的R区、S区和3'-端稍长且更多样化的R区[20](图1-C)。在超嗜热菌强烈炽热球菌(Pyrococcusfuriosus)中,Cas6的功能与CasE相同,Cas6是金属离子依赖型的核酸内切酶,与CRISPR的重复序列5'-端特定基序结合并在重复序列的3'-端将其切开[21]。CRISPR簇及cas基因的转录受到宿主内多种因素的调控[22]。在大肠杆菌中,类组蛋白拟核构造蛋白(histone-likenucleoidstructuringprotein,H-NS)能降低CRISPR系统抵抗噬菌体的能力[23],而转录因子LeuO能解除H-NS的抑制作用[24]。研究表明,H-NS能与CRISPR簇及cas基因的启动子区结合,抑制CRISPR簇及cas基因的转录(图1-C),一方面,CRISPR簇被转录的前crRNA减少了,另一方面,CAS蛋白特别是CasE的减少又导致将半衰期短的前crRNA加工成更稳定的crRNA的能力降低[23],从而导致CRISPR系统抵抗噬菌体的能力降低。4.3CRISPR系统介导的沉默嗜热链球菌在受噬菌体侵染后,CRISPR簇新获得的间区序列有的来自噬菌体DNA的编码链,有的来自模板链[7];无论用λ噬菌体的编码链还是用模板链作为人工设计的间区序列,都能有效地使大肠杆菌获得抗λ噬菌体的能力[20];表皮葡萄球菌的1个CRISPR间区序列与结合转移质粒的切口酶(nickase)具有同源性,而切口酶只在结合转移的供体菌中发挥功能,但CRISPR系统能使供体菌和受体菌都失去结合转移能力[8]。综合这些研究结果,不难看出crRNA的作用机理不同于反义RNA,应该是直接作用于DNA。同时,前间区序列邻近基序PAM不仅是选取新间区序列的识别位点,也是crRNA作用于外来DNA的靶标之一[4]。强烈炽热球菌的Cascade复合物包括多个Cmr型的CAS蛋白,由Cas6加工成的前导RNA与该复合物结合后继续被加工,即得到成熟的psiRNA(prokaryoticsilencingRNA)[25]。Hale等进行的体外实验表明,单链的psiRNA-Cmr复合物能位点特异地切割RNA而不能作用于DNA,而且切割位点总位于psiRNA的3'-末端往前14nt。这种作用机理与真核生物的siRNA极其相似,只是还没有psiRNA作用于天然靶标(来自噬菌体、质粒的转录体)的体内证据[26]。因此,前crRNA加工为成熟的crRNA后,是直接作用于DNA还是RNA,至今仍处于争论期,或许二者兼而有之,但谁主谁次尚不清楚。4.4自我与非我的区分一般情况下,免疫系统能很好地区分自我与非我,从而快速产生针对外来成分的免疫反应,同时避免产生自身免疫。Marraffini等通过向敲除了CRISPR簇的表皮葡萄球菌菌株中回补不同的CRISPR簇的突变体,并转入带有前间区序列及其邻近基序PAM(进行了个别重要碱基的替换)的外来DNA序列,他们发现成熟crRNA的5'-端重复序列(R区)在保护自身CRISPR簇不被切割时起主要作用,也暗示成熟crRNA5'-端的均质性(5'-端R区为8-9nt,3'-端R区更长且多变)对crRNA发挥功能的重要性(图2);同时,外来DNA中的前间区序列邻近基序PAM在区分自我与非我的过程中也起关键作用。值得关注的是,Stern等最近发现了自靶向的间区序列(self-targetingspacers)。他们分析了来自330种含CRISPR系统的原核生物的23550个间区序列(S区),发现每250个间区序列中就有1个是自靶向的,所研究的330种原核生物中有18%存在自靶向的间区序列。由于缺乏保守性,而且在自靶向间区序列的附近存在大量已退化的重复序列,因此,他们认为存在自靶向的间区序列不是其他调控机制而是一种自身免疫机制[28]。图2crRNA区分自我与非我的机制5展望综上所述,深入研




 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码










 灵蝶儿
灵蝶儿
本文标题:CRISPR及其在原核生物防御系统中作用的研究进展
链接地址:https://www.777doc.com/doc-2907024 .html